Medicinsk ekspert af artiklen

Nye publikationer
Syntese, udskillelse og metabolisme af katekolaminer
Sidst revideret: 06.07.2025

Alt iLive-indhold gennemgås medie eller kontrolleres for at sikre så meget faktuel nøjagtighed som muligt.
Vi har strenge sourcing retningslinjer og kun link til velrenommerede medie websteder, akademiske forskningsinstitutioner og, når det er muligt, medicinsk peer reviewed undersøgelser. Bemærk at tallene inden for parentes ([1], [2] osv.) Er klikbare links til disse undersøgelser.
Hvis du mener, at noget af vores indhold er unøjagtigt, forældet eller på anden måde tvivlsomt, skal du vælge det og trykke på Ctrl + Enter.
Binyremarven producerer forbindelser med en struktur, der langt fra er steroider. De indeholder en 3,4-dihydroxyphenyl (catechol)-kerne og kaldes katekolaminer. Disse omfatter adrenalin, noradrenalin og dopamin beta-hydroxytyramin.
Sekvensen af katekolaminsyntese er ret simpel: tyrosin → dihydroxyphenylalanin (DOPA) → dopamin → noradrenalin → adrenalin. Tyrosin kommer ind i kroppen med mad, men kan også dannes fra phenylalanin i leveren under påvirkning af phenylalaninhydroxylase. Slutprodukterne af tyrosinomdannelse i væv er forskellige. I binyremarven fortsætter processen til stadiet med dannelse af adrenalin, i enderne af de sympatiske nerver - noradrenalin, i nogle neuroner i centralnervesystemet slutter syntesen af katekolaminer med dannelsen af dopamin.
Omdannelsen af tyrosin til DOPA katalyseres af tyrosinhydroxylase, hvis kofaktorer er tetrahydrobiopterin og oxygen. Det menes, at dette enzym begrænser hastigheden af hele processen med katekolaminbiosyntese og hæmmes af processens slutprodukter. Tyrosinhydroxylase er det primære mål for regulatoriske effekter på katekolaminbiosyntese.
Omdannelsen af DOPA til dopamin katalyseres af enzymet DOPA-decarboxylase (cofaktor - pyridoxalphosphat), som er relativt uspecifik og decarboxylerer andre aromatiske L-aminosyrer. Der er dog indikationer på muligheden for at modificere syntesen af katekolaminer ved at ændre aktiviteten af dette enzym. Nogle neuroner mangler enzymer til den videre omdannelse af dopamin, og det er slutproduktet. Andre væv indeholder dopamin beta-hydroxylase (cofaktorer - kobber, ascorbinsyre og ilt), som omdanner dopamin til noradrenalin. I binyremarven (men ikke i enderne af de sympatiske nerver) er phenylethanolamin til stede - en methyltransferase, der danner adrenalin fra noradrenalin. I dette tilfælde fungerer S-adenosylmethionin som donor af methylgrupper.
Det er vigtigt at huske, at syntesen af phenylethanolamin-N-methyltransferase induceres af glukokortikoider, der kommer ind i medulla fra cortex via det portale venesystem. Dette kan forklare, at to forskellige endokrine kirtler er kombineret i ét organ. Betydningen af glukokortikoider for syntesen af adrenalin understreges af, at cellerne i binyremarven, der producerer noradrenalin, er placeret omkring arteriekarrene, hvorimod de adrenalinproducerende celler hovedsageligt modtager blod fra de venøse bihuler, der er placeret i binyrebarken.
Nedbrydningen af katekolaminer sker hovedsageligt under påvirkning af to enzymsystemer: catechol-O-methyltransferase (COMT) og monoaminoxidase (MAO). De vigtigste nedbrydningsveje for adrenalin og noradrenalin er skematisk vist i figur 54. Under påvirkning af COMT i nærvær af methylgruppedonoren S-adrenosylmethionin omdannes katekolaminer til normetanephrin og metanephrin (3-O-methylderivater af noradrenalin og adrenalin), som under påvirkning af MAO omdannes til aldehyder og derefter (i nærvær af aldehydoxidase) til vanillylmandelsyre (VMA), det primære nedbrydningsprodukt af noradrenalin og adrenalin. I samme tilfælde, når katekolaminer initialt udsættes for MAO i stedet for COMT, omdannes de til 3,4-dioxomandelaldehyd og derefter, under påvirkning af aldehydoxidase og COMT, til 3,4-dioxomandelsyre og VMC. I nærvær af alkoholdehydrogenase kan 3-methoxy-4-oxyphenylglycol, som er det primære slutprodukt af adrenalin- og noradrenalin-nedbrydning i centralnervesystemet, dannes fra katekolaminer.
Nedbrydningen af dopamin er lignende, bortset fra at dens metabolitter mangler hydroxylgruppen ved beta-kulstofatomet, og derfor dannes der homovanillinsyre (HVA) eller 3-methoxy-4-hydroxyphenyleddikesyre i stedet for vanillylmandelsyre.
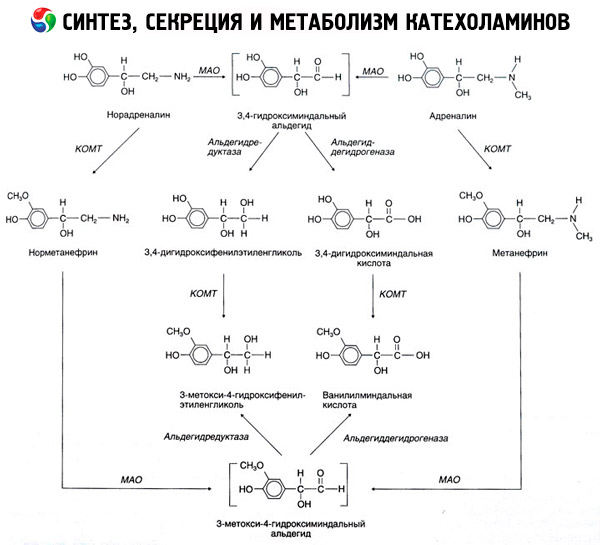
Eksistensen af en quinoid oxidationsvej for katekolaminmolekylet, som kan producere mellemprodukter med udtalt biologisk aktivitet, postuleres også.
Noradrenalin og adrenalin, der dannes under påvirkning af cytosoliske enzymer i de sympatiske nerveender og binyremarven, trænger ind i de sekretoriske granuler, hvilket beskytter dem mod nedbrydningsenzymer. Granulernes optagelse af katekolaminer kræver energiforbrug. I binyremarvens kromaffingranuler er katekolaminer tæt bundet til ATP (i forholdet 4:1) og specifikke proteiner - kromograniner, hvilket forhindrer diffusion af hormoner fra granulerne ind i cytoplasmaet.
Den direkte stimulus for udskillelsen af katekolaminer er tilsyneladende calciums indtrængen i cellen, hvilket stimulerer exocytose (fusion af granulamembranen med celleoverfladen og deres brud med fuldstændig frigivelse af opløseligt indhold - katekolaminer, dopamin beta-hydroxylase, ATP og chromograniner - i den ekstracellulære væske).
Fysiologiske virkninger af katekolaminer og deres virkningsmekanisme
Virkningerne af katekolaminer begynder med interaktion med specifikke receptorer i målcellerne. Mens receptorer for skjoldbruskkirtel- og steroidhormoner er lokaliseret inde i cellerne, er receptorer for katekolaminer (såvel som acetylcholin og peptidhormoner) til stede på den ydre celleoverflade.
Det har længe været fastslået, at adrenalin eller noradrenalin i forhold til nogle reaktioner er mere effektive end den syntetiske katekolamin isoproterenol, mens isoproterenols effekt i forhold til andre er bedre end adrenalins eller noradrenalins virkninger. På dette grundlag blev der udviklet et koncept om tilstedeværelsen af to typer adrenoreceptorer i væv: alfa og beta, og i nogle af dem kan kun én af disse to typer være til stede. Isoproterenol er den kraftigste agonist af beta-adrenoreceptorer, mens den syntetiske forbindelse phenylephrin er den kraftigste agonist af alfa-adrenoreceptorer. Naturlige katekolaminer - adrenalin og noradrenalin - er i stand til at interagere med receptorer af begge typer, men adrenalin udviser en større affinitet for beta-, og noradrenalin - for alfa-receptorer.
Katekolaminer aktiverer kardiale beta-adrenerge receptorer kraftigere end glatmuskulatur beta-receptorer, hvilket gjorde det muligt at opdele beta-typen i undertyper: beta1-receptorer (hjerte, fedtceller) og beta2-receptorer (bronkier, blodkar osv.). Effekten af isoproterenol på beta1-receptorer overstiger effekten af adrenalin og noradrenalin kun 10 gange, mens den på beta2-receptorer virker 100-1000 gange stærkere end naturlige katekolaminer.
Brugen af specifikke antagonister (phentolamin og phenoxybenzamin for alfa- og propranolol for beta-receptorer) bekræftede tilstrækkeligheden af klassificeringen af adrenoreceptorer. Dopamin er i stand til at interagere med både alfa- og beta-receptorer, men forskellige væv (hjerne, hypofyse, kar) har også deres egne dopaminerge receptorer, hvis specifikke blokker er haloperidol. Antallet af beta-receptorer varierer fra 1000 til 2000 pr. celle. De biologiske virkninger af katekolaminer medieret af beta-receptorer er normalt forbundet med aktiveringen af adenylatcyklase og en stigning i det intracellulære indhold af cAMP. Selvom receptoren og enzymet er funktionelt forbundet, er de forskellige makromolekyler. Guanosintrifosfat (GTP) og andre purinnukleotider deltager i moduleringen af adenylatcyklaseaktivitet under påvirkning af hormon-receptorkomplekset. Ved at øge enzymaktiviteten synes de at reducere beta-receptorernes affinitet for agonister.
Fænomenet med øget følsomhed i denerverede strukturer har længe været kendt. Tværtimod reducerer langvarig eksponering for agonister følsomheden af målvæv. Undersøgelsen af beta-receptorer har gjort det muligt at forklare disse fænomener. Det er blevet vist, at langvarig eksponering for isoproterenol fører til et tab af følsomhed af adenylatcyklase på grund af et fald i antallet af beta-receptorer.
Desensibiliseringsprocessen kræver ikke aktivering af proteinsyntese og skyldes sandsynligvis den gradvise dannelse af irreversible hormon-receptorkomplekser. Tværtimod ledsages introduktionen af 6-oxidopamin, som ødelægger sympatiske ender, af en stigning i antallet af reagerende beta-receptorer i væv. Det er muligt, at en stigning i sympatisk nerveaktivitet også forårsager aldersrelateret desensibilisering af blodkar og fedtvæv i forhold til katekolaminer.
Antallet af adrenoreceptorer i forskellige organer kan kontrolleres af andre hormoner. Således øges østradiol, og progesteron mindsker antallet af alfa-adrenoreceptorer i livmoderen, hvilket ledsages af en tilsvarende stigning og fald i dens kontraktile respons på katekolaminer. Hvis den intracellulære "second messenger", der dannes ved virkningen af beta-receptoragonister, helt sikkert er cAMP, er situationen med hensyn til transmitteren af alfa-adrenerge effekter mere kompliceret. Eksistensen af forskellige mekanismer antages: et fald i niveauet af cAMP, en stigning i indholdet af cAMP, modulering af cellulær calciumdynamik osv.
For at reproducere forskellige effekter i kroppen kræves der normalt doser af adrenalin, der er 5-10 gange mindre end noradrenalin. Selvom sidstnævnte er mere effektivt i forhold til α- og beta1-adrenalinreceptorer, er det vigtigt at huske, at begge endogene katekolaminer er i stand til at interagere med både alfa- og beta-receptorer. Derfor afhænger et givet organs biologiske respons på adrenerg aktivering i høj grad af typen af receptorer, der er til stede i det. Dette betyder dog ikke, at selektiv aktivering af det nervøse eller humorale led i det sympatiske binyresystem er umulig. I de fleste tilfælde observeres øget aktivitet af dets forskellige led. Det er således generelt accepteret, at hypoglykæmi refleksivt aktiverer binyremarven, mens et fald i blodtrykket (postural hypotension) hovedsageligt ledsages af frigivelsen af noradrenalin fra enderne af de sympatiske nerver.
Adrenoreceptorer og virkningerne af deres aktivering i forskellige væv
System, organ |
Adrenerg receptortype |
Reaktion |
Kardiovaskulært system: |
||
Hjerte |
Beta |
Øget hjertefrekvens, ledningsevne og kontraktilitet |
Arterioler: |
||
Hud og slimhinder |
Alfa |
Reduktion |
Skeletmuskler |
Beta |
Ekspansionskontraktion |
Maveorganer |
Alfa (mere) |
Reduktion |
Beta |
Udvidelse |
|
Vener |
Alfa |
Reduktion |
Åndedrætssystem: |
||
Musklerne i bronkierne |
Beta |
Udvidelse |
Fordøjelsessystemet: |
||
Mave |
Beta |
Nedsatte motoriske færdigheder |
Tarme |
Alfa |
Sammentrækning af lukkemusklerne |
Milt |
Alfa |
Reduktion |
Beta |
Lempelse |
|
Eksokrin bugspytkirtel |
Alfa |
Nedsat sekretion |
Urogenitalsystemet: |
Alfa |
Sphincter-kontraktion |
Blære |
Beta |
Afslapning af ejektormusklen |
Mandlige kønsorganer |
Alfa |
Ejakulation |
Øjne |
Alfa |
Pupiludvidelse |
Læder |
Alfa |
Øget svedtendens |
Spytkirtler |
Alfa |
Udskillelse af kalium og vand |
Beta |
Amylase-sekretion |
|
Endokrine kirtler: |
||
Øer i bugspytkirtlen |
||
Betaceller |
Alfa (mere) |
Nedsat insulinudskillelse |
Beta |
Øget insulinudskillelse |
|
Alfaceller |
Beta |
Øget udskillelse af glukagon |
8-celler |
Beta |
Øget sekretion af somatostatin |
Hypothalamus og hypofyse: |
||
Somatotrofer |
Alfa |
Øget sekretion af STH |
Beta |
Nedsat sekretion af STH |
|
Laktotrofer |
Alfa |
Nedsat sekretion af prolaktin |
Thyrotrofer |
Alfa |
Nedsat TSH-sekretion |
Kortikotrofer |
Alfa |
Øget ACTH-sekretion |
| beta | Nedsat ACTH-sekretion | |
Skjoldbruskkirtel: |
||
Follikulære celler |
Alfa |
Nedsat udskillelse af thyroxin |
Beta |
Øget sekretion af thyroxin |
|
Parafollikulære (K) celler |
Beta |
Øget sekretion af calcitonin |
Biskjoldbruskkirtlerne |
Beta |
Øget sekretion af PTH |
Nyrer |
Beta |
Øget reninsekretion |
Mave |
Beta |
Øget sekretion af gastrin |
BX |
Beta |
Øget iltforbrug |
Lever |
? |
Øget glykogenolyse og glukoneogenese med frigivelse af glukose; øget ketogenese med frigivelse af ketonstoffer |
Fedtvæv |
Beta |
Øget lipolyse med frigivelse af frie fedtsyrer og glycerol |
Skeletmuskler |
Beta |
Øget glykolyse med frigivelse af pyruvat og laktat; nedsat proteolyse med et fald i frigivelsen af alanin, glutamin |
Det er vigtigt at tage højde for, at resultaterne af intravenøs administration af katekolaminer ikke altid afspejler virkningerne af endogene forbindelser tilstrækkeligt. Dette gælder primært for noradrenalin, da det i kroppen hovedsageligt ikke frigives i blodet, men direkte i de synaptiske kløfter. Derfor aktiverer endogen noradrenalin for eksempel ikke kun vaskulære alfa-receptorer (forhøjet blodtryk), men også beta-receptorer i hjertet (øget hjertefrekvens), hvorimod tilførsel af noradrenalin udefra primært fører til aktivering af vaskulære alfa-receptorer og en refleks (via vagus) nedsættelse af hjerteslaget.
Lave doser adrenalin aktiverer primært beta-receptorer i muskelkarrene og hjertet, hvilket resulterer i et fald i den perifere vaskulære modstand og en stigning i hjertets minutvolumen. I nogle tilfælde kan den første effekt være fremherskende, og hypotension udvikles efter administration af adrenalin. I højere doser aktiverer adrenalin også alfa-receptorer, hvilket ledsages af en stigning i den perifere vaskulære modstand og, på baggrund af en stigning i hjertets minutvolumen, fører til en stigning i arterielt tryk. Imidlertid bevares dens effekt på vaskulære beta-receptorer også. Som følge heraf overstiger stigningen i systolisk tryk den tilsvarende indikator for diastolisk tryk (stigning i pulstryk). Ved introduktion af endnu højere doser begynder adrenalinens alfa-mimetiske virkninger at være fremherskende: systolisk og diastolisk tryk stiger parallelt, som under påvirkning af noradrenalin.
Katekolaminers effekt på stofskiftet består af deres direkte og indirekte effekter. Førstnævnte realiseres hovedsageligt gennem beta-receptorer. Mere komplekse processer er forbundet med leveren. Selvom øget leverglykogenolyse traditionelt anses for at være et resultat af beta-receptoraktivering, er der også tegn på alfa-receptorinvolvering. Indirekte effekter af katekolaminer er forbundet med modulering af sekretionen af mange andre hormoner, såsom insulin. I adrenalins effekt på dets sekretion dominerer den alfa-adrenerge komponent klart, da det har vist sig, at enhver stress ledsages af hæmning af insulinsekretion.
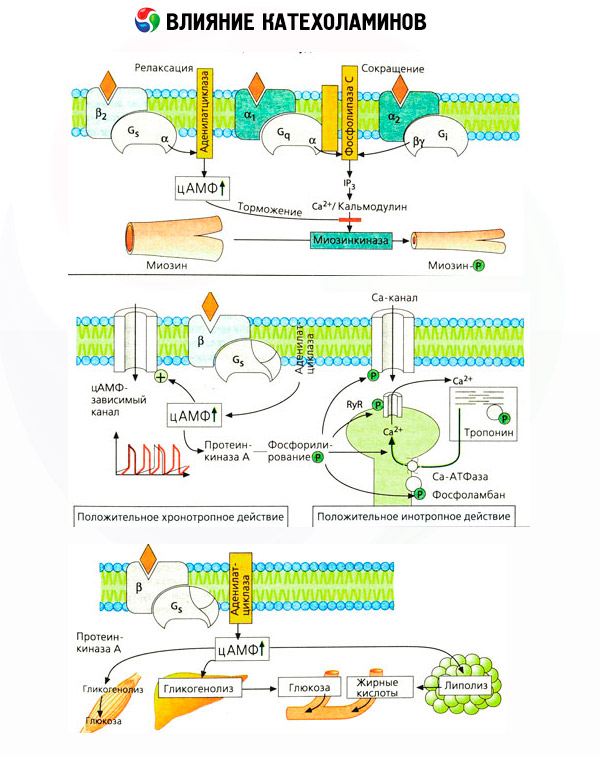
Kombinationen af direkte og indirekte virkninger af katekolaminer forårsager hyperglykæmi, som ikke kun er forbundet med øget hepatisk glukoseproduktion, men også med hæmning af dens udnyttelse i perifert væv. Acceleration af lipolyse forårsager hyperlipacidæmi med øget tilførsel af fedtsyrer til leveren og intensivering af ketonstofproduktion. Øget glykolyse i muskler fører til en øget frigivelse af laktat og pyruvat i blodet, som sammen med glycerol frigivet fra fedtvæv tjener som forløbere for hepatisk glukoneogenese.
Regulering af katekolaminsekretion. Ligheden mellem produkterne og reaktionsmetoderne i det sympatiske nervesystem og binyremarven var grundlaget for at kombinere disse strukturer til et enkelt sympatisk-binyresystem i kroppen med allokering af dets nervøse og hormonelle forbindelser. Forskellige afferente signaler er koncentreret i hypothalamus og centrene i rygmarven og medulla oblongata, hvorfra efferente signaler stammer og skifter til cellelegemerne af præganglioniske neuroner placeret i rygmarvens laterale horn på niveau med VIII cervikale - II-III lumbale segmenter.
De præganglionære axoner i disse celler forlader rygmarven og danner synaptiske forbindelser med neuroner placeret i ganglierne i den sympatiske kæde eller med celler i binyremarven. Disse præganglionære fibre er kolinerge. Den første fundamentale forskel mellem sympatiske postganglionære neuroner og kromaffinceller i binyremarven er, at sidstnævnte transmitterer det kolinerge signal, de modtager, ikke via nerveledning (postganglionære adrenerge nerver), men via den humorale bane, hvorved adrenerge forbindelser frigives til blodet. Den anden forskel er, at postganglionære nerver producerer noradrenalin, mens celler i binyremarven primært producerer adrenalin. Disse to stoffer har forskellige virkninger på væv.