Medicinsk ekspert af artiklen

Nye publikationer
Hypofysen
Sidst revideret: 04.07.2025

Alt iLive-indhold gennemgås medie eller kontrolleres for at sikre så meget faktuel nøjagtighed som muligt.
Vi har strenge sourcing retningslinjer og kun link til velrenommerede medie websteder, akademiske forskningsinstitutioner og, når det er muligt, medicinsk peer reviewed undersøgelser. Bemærk at tallene inden for parentes ([1], [2] osv.) Er klikbare links til disse undersøgelser.
Hvis du mener, at noget af vores indhold er unøjagtigt, forældet eller på anden måde tvivlsomt, skal du vælge det og trykke på Ctrl + Enter.
Hypofysen (hypofysen, s.glandula pituitaria) er placeret i hypofysens fossa i kilebenets sella turcica og er adskilt fra kraniehulen ved en processus dura mater i hjernen, der danner sellas diafragma. Gennem åbningen i dette diafragma er hypofysen forbundet med infundibulum i hypothalamus i diencephalon. Hypofysens tværgående størrelse er 10-17 mm, den anteroposterior er 5-15 mm, den vertikale er 5-10 mm. Hypofysens masse hos mænd er cirka 0,5 g, hos kvinder - 0,6 g. Hypofysen er udvendigt dækket af en kapsel.
I overensstemmelse med hypofysens udvikling fra to forskellige grundstoffer skelnes der i organet mellem to lapper - den forreste og den bageste. Adenohypofysen, eller forlappen (adenohypophysis, s.lobus anterior), er større og udgør 70-80% af hypofysens samlede masse. Den er tættere end den bageste lap. I den forreste lap skelnes en distal del (pars distalis), som optager den forreste del af hypofysefossaen, en mellemliggende del (pars intermedia), der er placeret på grænsen til den bageste lap, og en knolddel (pars tuberalis), der går opad og forbinder med hypothalamus' infundibulum. På grund af overfloden af blodkar har den forreste lap en lysegul farve med et rødligt skær. Parenkym i den forreste hypofyse er repræsenteret af flere typer kirtelceller, mellem hvis strenge der er placeret sinusformede blodkapillærer. Halvdelen (50%) af adenohypofysecellerne er kromofile adenocytter, som har finkornede granuler i deres cytoplasma, der farver godt med kromsalte. Disse er acidofile adenocytter (40% af alle adenohypofyseceller) og basofile adenocytter (10%). Basofile adenocytter omfatter gonadotrope, kortikotrope og thyrotrope endokrinocytter. Kromofobe adenocytter er små, de har en stor cellekerne og en lille mængde cytoplasma. Disse celler betragtes som forstadier til kromofile adenocytter. De andre 50% af adenohypofysecellerne er kromofobe adenocytter.
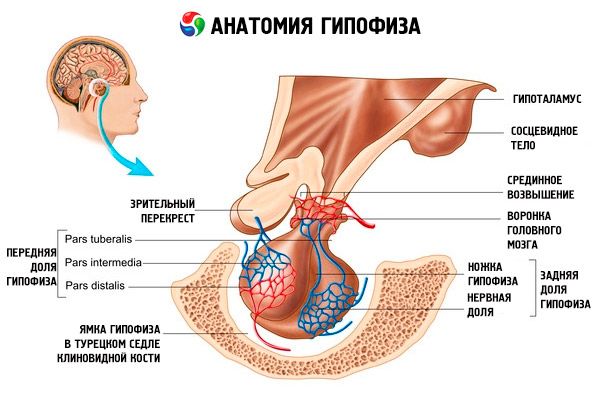
Neurohypofysen, eller den bageste lap (neurohypophysis, s.lobus posterior), består af neurallappen (lobus nervosus), som er placeret i den bageste del af hypofysen, og tragten (infundibulum), der er placeret bag den knolde del af adenohypofysen. Hypofysens bageste lap dannes af neurogliaceller (hypofyseceller), nervefibre, der går fra hypothalamus' neurosekretoriske kerner til neurohypofysen, og neurosekretoriske blodlegemer.
Hypofysen er, via nervefibre (baner) og blodkar, funktionelt forbundet med hypothalamus i diencephalon, som regulerer hypofysens aktivitet. Hypofysen og hypothalamus betragtes sammen med deres neuroendokrine, vaskulære og nerveforbindelser normalt som det hypothalamus-hypofysesystem.
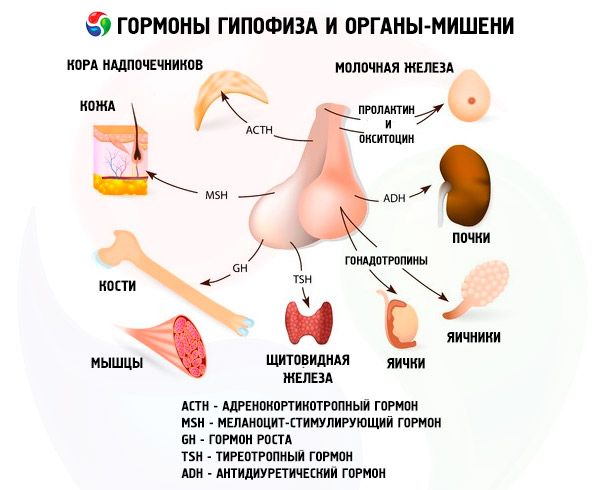
Hormoner i den forreste og bageste hypofyse påvirker mange af kroppens funktioner, primært gennem andre endokrine kirtler. I den forreste hypofyse producerer acidofile adenocytter (alfaceller) somatotropisk hormon (væksthormon), som deltager i reguleringen af vækst- og udviklingsprocesser hos unge organismer. Kortikotrope endokrinocytter udskiller adrenokortikotropisk hormon (ACTH), som stimulerer binyrernes udskillelse af steroidhormoner. Thyrotrope endokrinocytter udskiller thyrotropisk hormon (TSH), som påvirker udviklingen af skjoldbruskkirtlen og aktiverer produktionen af dens hormoner. Gonadotrope hormoner: follikelstimulerende (FSH), luteiniserende (LH) og prolaktin - påvirker kroppens seksuelle modning, regulerer og stimulerer udviklingen af follikler i æggestokkene, ægløsning, vækst af mælkekirtlerne og mælkeproduktion hos kvinder, samt spermatogenese hos mænd. Disse hormoner produceres af basofile adenocytter (betaceller). Lipotrope faktorer fra hypofysen udskilles også her, som påvirker mobiliseringen og udnyttelsen af fedtstoffer i kroppen. I den mellemliggende del af forlappen dannes melanocytstimulerende hormon, som styrer dannelsen af pigmenter - melaniner - i kroppen.
Neurosekretoriske celler i de supraoptiske og paraventrikulære kerner i hypothalamus producerer vasopressin og oxytocin. Disse hormoner transporteres til cellerne i den bageste hypofyse langs axoner, der udgør hypothalamus-hypofysen. Fra den bageste hypofyse kommer disse stoffer ind i blodbanen. Hormonet vasopressin har en vasokonstriktiv og antidiuretisk effekt, hvilket det også kaldes antidiuretisk hormon (ADH). Oxytocin har en stimulerende effekt på livmodermusklernes kontraktilitet, øger mælkesekretionen fra den ammende mælkekirtle, hæmmer udviklingen og funktionen af det gule legeme og påvirker ændringer i tonus i de glatte (ikke-stribede) muskler i mave-tarmkanalen.
Udvikling af hypofysen
Hypofysens forlap udvikler sig fra epitelet i mundhulens dorsale væg i form af en ringformet udvækst (Rathkes pose). Denne ektodermale fremspring vokser mod bunden af den fremtidige tredje ventrikel. Mod den, fra den nedre overflade af den anden hjerneblære (den fremtidige bund af den tredje ventrikel), vokser en proces, hvorfra infundibulums grå tuberkel og hypofysens baglap udvikler sig.
Hypofysens kar og nerver
Fra de indre halspulsårer og karrene i storhjernens arteriekreds ledes de øvre og nedre hypofysearterier til hypofysen. De øvre hypofysearterier går til den grå kerne og infundibulum i hypothalamus, anastomoserer med hinanden her og danner kapillærer, der trænger ind i hjernevævet - det primære hæmokapillære netværk. Fra de lange og korte løkker i dette netværk dannes portalvenerne, som ledes til hypofysens forlap. I parenkymet af hypofysens forlap opdeles disse vener i brede sinusformede kapillærer, der danner det sekundære hæmokapillære netværk. Hypofysens baglap forsynes hovedsageligt med blod fra den nedre hypofysearterie. Der er lange arterielle anastomoser mellem de øvre og nedre hypofysearterier. Udstrømningen af venøst blod fra det sekundære hæmokapillære netværk udføres gennem et system af vener, der strømmer ind i de kavernøse og interkavernøse bihuler i hjernens dura mater.
Sympatiske fibre, der kommer ind i organet sammen med arterierne, deltager i hypofysens innervation. Postganglioniske sympatiske nervefibre udgår fra plexus i den indre halspulsåre. Derudover findes der talrige ender af processerne i neurosekretoriske celler, der er placeret i hypothalamus' kerner, i hypofysens bageste lap.
Aldersrelaterede træk ved hypofysen
Hypofysens gennemsnitsvægt hos nyfødte når 0,12 g. Organets vægt fordobles ved 10-årsalderen og tredobles ved 15-årsalderen. Ved 20-årsalderen når hypofysens vægt sit maksimum (530-560 mg) og forbliver næsten uændret i efterfølgende aldersperioder. Efter 60 år observeres et lille fald i vægten af denne endokrine kirtel.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hypofysehormoner
Enheden af nervøs og hormonel regulering i kroppen sikres af den tætte anatomiske og funktionelle forbindelse mellem hypofysen og hypothalamus. Dette kompleks bestemmer tilstanden og funktionen af hele det endokrine system.
Den primære endokrine kirtel, der producerer en række peptidhormoner, som direkte regulerer funktionen af de perifere kirtler, er hypofysen. Det er en rødlig-grå bønneformet formation dækket af en fibrøs kapsel, der vejer 0,5-0,6 g. Den varierer lidt afhængigt af en persons køn og alder. Den generelt accepterede opdeling af hypofysen i to lapper, der adskiller sig i udvikling, struktur og funktion, forbliver: den forreste distale - adenohypofysen og den bageste - neurohypofysen. Den første udgør omkring 70% af kirtelens samlede masse og er traditionelt opdelt i den distale, infundibulære og mellemliggende del, den anden - i den bageste del, eller lap, og hypofysestilken. Kirtlen er placeret i hypofysefossaen i sella turcica i sphenoidbenet og er forbundet med hjernen gennem stilken. Den øverste del af den forreste lap er dækket af den optiske chiasme og de optiske kanaler. Blodforsyningen til hypofysen er meget rigelig og varetages af grene af den indre halspulsåre (hypofysens øvre og nedre arterier) samt af grene af hjernens arteriekreds. De øvre hypofysearterier deltager i blodforsyningen til adenohypofysen, og de nedre - neurohypofysen, i kontakt med de neurosekretoriske ender af axonerne i hypothalamus' store cellekerner. Førstnævnte går ind i hypothalamus' mediane eminens, hvor de spredes i et kapillærnetværk (den primære kapillærplexus). Disse kapillærer (som terminalerne af axonerne i de små neurosekretoriske celler i den mediobasale hypothalamus er i kontakt med) samles i portalvener, der nedstiger langs hypofysens stilk ind i adenohypofysens parenkym, hvor de igen deler sig i et netværk af sinusformede kapillærer (den sekundære kapillærplexus). Således kommer blodet, efter at have passeret gennem hypothalamus' mediane eminens, hvor det er beriget med hypothalamiske adenohypofysotrope hormoner (frigivende hormoner), ind i adenohypofysen.
Udstrømningen af blod mættet med adenohypofysehormoner fra talrige kapillærer i den sekundære plexus udføres gennem venesystemet, som igen strømmer ind i dura materens venøse bihuler og derefter ind i den generelle blodbane. Således er hypofysens portalsystem med en nedadgående retning af blodgennemstrømningen fra hypothalamus en morfofunktionel komponent i den komplekse mekanisme for neurohumoral kontrol af adenohypofysens tropiske funktioner.
Hypofysen er innerveret af sympatiske fibre, der følger hypofysearterierne. De udgår fra postganglionære fibre, der passerer gennem den indre carotis plexus, forbundet med de øvre cervikale ganglier. Der er ingen direkte innervation af adenohypofysen fra hypothalamus. Den bageste lap modtager nervefibre fra hypothalamus' neurosekretoriske kerner.
Adenohypofysen er en meget kompleks formation i sin histologiske arkitektur. Den har to typer kirtelceller - kromofobe og kromofile. Sidstnævnte er igen opdelt i acidofile og basofile (en detaljeret histologisk beskrivelse af hypofysen findes i det tilsvarende afsnit af manualen). Det skal dog bemærkes, at de hormoner, der produceres af de kirtelceller, der udgør adenohypofysens parenkym, på grund af sidstnævntes diversitet til en vis grad er forskellige i deres kemiske natur, og den fine struktur af de sekreterende celler skal svare til biosynteseegenskaberne hos hver af dem. Men nogle gange kan overgangsformer af kirtelceller, der er i stand til at producere flere hormoner, observeres i adenohypofysen. Der er tegn på, at typen af kirtelceller i adenohypofysen ikke altid er genetisk bestemt.
Under diafragmaet på sella turcica sidder den tragtformede del af forlappen. Den omslutter hypofysens stilk og har kontakt med den grå tuberkel. Denne del af adenohypofysen er karakteriseret ved tilstedeværelsen af epitelceller og rigelig blodforsyning. Den er også hormonelt aktiv.
Den mellemliggende (midterste) del af hypofysen består af flere lag af store sekretorisk aktive basofile celler.
Hypofysen udfører forskellige funktioner gennem sine hormoner. Dens forlap producerer adrenokortikotrope (ACTH), thyreoideastimulerende (TSH), follikelstimulerende (FSH), luteiniserende (LH), lipotrope hormoner, samt væksthormon - somatotropisk (STO) og prolaktin. I den mellemste lap syntetiseres melanocytstimulerende hormon (MSH), og i den bageste lap akkumuleres vasopressin og oxytocin.
ACTH
Hypofysehormoner er en gruppe af protein- og peptidhormoner og glykoproteiner. Af hormonerne i den forreste hypofyse er ACTH blevet undersøgt bedst. Det produceres af basofile celler. Dets primære fysiologiske funktion er at stimulere biosyntesen og sekretionen af steroidhormoner fra binyrebarken. ACTH udviser også melanocytstimulerende og lipotropisk aktivitet. I 1953 blev det isoleret i ren form. Senere blev dets kemiske struktur etableret, bestående af 39 aminosyrerester hos mennesker og en række pattedyr. ACTH har ingen artsspecificitet. I øjeblikket er der udført kemisk syntese af både selve hormonet og forskellige fragmenter af dets molekyle, der er mere aktive end naturlige hormoner. Hormonets struktur har to sektioner af peptidkæden, hvoraf den ene sikrer detektion og binding af ACTH til receptoren, og den anden giver en biologisk effekt. Det binder tilsyneladende til ACTH-receptoren gennem interaktionen mellem hormonets og receptorens elektriske ladninger. Rollen som den biologiske effektor af ACTH udføres af fragmentet af 4-10-molekylet (Met-Glu-His-Phen-Arg-Tri-Tri).
ACTHs melanocytstimulerende aktivitet skyldes tilstedeværelsen i molekylet af den N-terminale region, der består af 13 aminosyrerester og gentager strukturen af alfa-melanocytstimulerende hormon. Denne region indeholder også et heptapeptid, der findes i andre hypofysehormoner, og som besidder en vis adrenokortikotropisk, melanocytstimulerende og lipotropisk aktivitet.
Det afgørende øjeblik i ACTHs virkning anses for at være aktiveringen af enzymet proteinkinase i cytoplasmaet med deltagelse af cAMP. Fosforyleret proteinkinase aktiverer enzymet esterase, som omdanner kolesterolestere til et frit stof i fedtdråber. Proteinet, der syntetiseres i cytoplasmaet som følge af fosforylering af ribosomer, stimulerer bindingen af frit kolesterol til cytokrom P-450 og dets overførsel fra lipiddråber til mitokondrier, hvor alle de enzymer, der sikrer omdannelsen af kolesterol til kortikosteroider, er til stede.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Thyroid-stimulerende hormon
TSH - thyrotropin - den primære regulator for udvikling og funktion af skjoldbruskkirtlen, samt processerne for syntese og sekretion af skjoldbruskkirtelhormoner. Dette komplekse protein - glykoprotein - består af alfa- og beta-underenheder. Strukturen af den første underenhed svarer til alfa-underenheden af luteiniserende hormon. Desuden er den stort set den samme hos forskellige dyrearter. Sekvensen af aminosyrerester i beta-underenheden af human TSH er blevet dechifreret og består af 119 aminosyrerester. Det kan bemærkes, at beta-underenhederne af human og kvæg-TSH er ens på mange måder. De biologiske egenskaber og arten af den biologiske aktivitet af glykoproteinhormoner bestemmes af beta-underenheden. Den sikrer også hormonets interaktion med receptorer i forskellige målorganer. Beta-underenheden udviser dog hos de fleste dyr kun specifik aktivitet, efter at den kombineres med alfa-underenheden, som fungerer som en slags aktivator af hormonet. Sidstnævnte inducerer med lige stor sandsynlighed luteiniserende, follikelstimulerende og thyrotropiske aktiviteter, der bestemmes af beta-underenhedens egenskaber. Den fundne lighed tillader os at konkludere, at disse hormoner opstod i evolutionsprocessen fra en fælles forgænger, hvor beta-underenheden også bestemmer hormonernes immunologiske egenskaber. Der er en antagelse om, at alfa-underenheden beskytter beta-underenheden mod virkningen af proteolytiske enzymer og også letter dens transport fra hypofysen til de perifere målorganer.
Gonadotrope hormoner
Gonadotropiner findes i kroppen i form af LH og FSH. Disse hormoners funktionelle formål er generelt reduceret til at sikre reproduktionsprocesser hos individer af begge køn. De er, ligesom TSH, komplekse proteiner - glykoproteiner. FSH inducerer modningen af follikler i kvinders æggestokke og stimulerer spermatogenesen hos mænd. LH forårsager follikelruptur hos kvinder med dannelsen af corpus luteum og stimulerer udskillelsen af østrogener og progesteron. Hos mænd accelererer det samme hormon udviklingen af interstitielt væv og udskillelsen af androgener. Gonadotropinernes virkninger er afhængige af hinanden og forekommer synkront.
Dynamikken i gonadotropinsekretionen hos kvinder ændrer sig i løbet af menstruationscyklussen og er blevet undersøgt tilstrækkeligt detaljeret. I den præovulatoriske (follikulære) fase af cyklussen er LH-indholdet på et forholdsvis lavt niveau, mens FSH er forhøjet. Efterhånden som folliklen modnes, øges østradiolsekretionen, hvilket fremmer øget produktion af gonadotropiner fra hypofysen og forekomsten af både LH- og FSH-cyklusser, dvs. kønshormoner stimulerer sekretionen af gonadotropiner.
I øjeblikket er strukturen af LH blevet bestemt. Ligesom TSH består den af 2 underenheder: a og b. Strukturen af alfa-underenheden af LH i forskellige dyrearter er stort set sammenfaldende, den svarer til strukturen af alfa-underenheden af TSH.
Strukturen af beta-underenheden i LH adskiller sig markant fra strukturen af beta-underenheden i TSH, selvom den har fire identiske sektioner af peptidkæden bestående af 4-5 aminosyrerester. I TSH er de lokaliseret i positionerne 27-31, 51-54, 65-68 og 78-83. Da beta-underenheden i LH og TSH bestemmer hormonernes specifikke biologiske aktivitet, kan det antages, at homologe sektioner i LH's og TSH's struktur bør sikre forbindelsen mellem beta-underenhederne og alfa-underenheden, og sektioner, der adskiller sig i struktur, bør være ansvarlige for specificiteten af hormonernes biologiske aktivitet.
Naturligt LH er meget stabilt over for proteolytiske enzymers virkning, men beta-underenheden spaltes hurtigt af chymotrypsin, og a-underenheden er vanskelig at hydrolysere af enzymet, dvs. den spiller en beskyttende rolle og forhindrer chymotrypsin i at få adgang til peptidbindingerne.
Hvad angår den kemiske struktur af FSH, har forskerne endnu ikke opnået endelige resultater. Ligesom LH består FSH af to underenheder, men beta-underenheden af FSH adskiller sig fra beta-underenheden af LH.
Prolaktin
Et andet hormon, prolaktin (laktogent hormon), spiller en aktiv rolle i reproduktionsprocesserne. De vigtigste fysiologiske egenskaber ved prolaktin hos pattedyr manifesterer sig i form af stimulering af udviklingen af mælkekirtler og amning, vækst af talgkirtler og indre organer. Det fremmer manifestationen af steroiders effekt på sekundære seksuelle karakteristika hos hanner, stimulerer den sekretoriske aktivitet af corpus luteum hos mus og rotter og deltager i reguleringen af fedtstofskiftet. Der er i de senere år blevet lagt stor vægt på prolaktin som en regulator af moderens adfærd; en sådan polyfunktionalitet forklares af dets evolutionære udvikling. Det er et af de gamle hypofysehormoner og findes endda hos padder. I øjeblikket er prolaktins struktur hos nogle pattedyrarter blevet fuldstændig afdækket. Indtil for nylig udtrykte forskere dog tvivl om eksistensen af et sådant hormon hos mennesker. Mange troede, at dets funktion blev udført af væksthormonet. Nu er der opnået overbevisende beviser for tilstedeværelsen af prolaktin hos mennesker, og dets struktur er delvist afdækket. Prolaktinreceptorer binder aktivt væksthormon og placentalaktogen, hvilket indikerer en enkelt virkningsmekanisme for de tre hormoner.
Somatotropin
Væksthormonet somatotropin har et endnu bredere virkningsspektrum end prolaktin. Ligesom prolaktin produceres det af acidofile celler i adenohypofysen. Somatotropin stimulerer skeletvækst, aktiverer proteinbiosyntese, har en fedtmobiliserende effekt og fremmer en forøgelse af kropsstørrelse. Derudover koordinerer det metaboliske processer.
Hormonets deltagelse i sidstnævnte bekræftes af en kraftig stigning i dets udskillelse fra hypofysen, for eksempel når blodsukkerniveauet falder.
Den kemiske struktur af dette humane hormon er nu fuldt etableret - 191 aminosyrerester. Dets primære struktur ligner den af chorionisk somatomammotropin eller placentalaktogen. Disse data indikerer en betydelig evolutionær nærhed mellem de to hormoner, selvom de udviser forskelle i biologisk aktivitet.
Det er nødvendigt at understrege den høje artsspecificitet af det pågældende hormon - for eksempel er STH af animalsk oprindelse inaktivt hos mennesker. Dette forklares både af reaktionen mellem STH-receptorerne hos mennesker og dyr og selve hormonets struktur. I øjeblikket er der undersøgelser i gang for at identificere aktive centre i STH's komplekse struktur, der udviser biologisk aktivitet. Individuelle fragmenter af molekylet, der udviser andre egenskaber, undersøges. For eksempel blev et peptid bestående af 14 aminosyrerester og svarende til molekyleafsnittet 31-44 isoleret efter hydrolyse af humant STH med pepsin. Det havde ingen væksteffekt, men overgik signifikant det native hormons lipotropiske aktivitet. Humant væksthormon har, i modsætning til lignende hormoner hos dyr, betydelig laktogen aktivitet.
Adenohypofysen syntetiserer mange peptid- og proteinstoffer, der har en fedtmobiliserende effekt, og de hypofysetropiske hormoner - ACTH, STH, TSH og andre - har en lipotropisk effekt. I de senere år er der blevet lagt særlig vægt på beta- og y-lipotropiske hormoner (LPG). De biologiske egenskaber ved beta-LPG er blevet undersøgt meget detaljeret; udover lipotropisk aktivitet har det også en melanocytstimulerende, kortikotropinstimulerende og hypocalcæmisk effekt og producerer også en insulinlignende effekt.
I øjeblikket er den primære struktur af fåre-LPG (90 aminosyrerester), lipotropiske hormoner fra svin og kvæg, blevet afkodet. Dette hormon har artsspecificitet, selvom strukturen af den centrale region af beta-LPG er den samme i forskellige arter. Det bestemmer hormonets biologiske egenskaber. Et af fragmenterne af denne region findes i strukturen af alfa-MSH, beta-MSH, ACTH og beta-LPG. Det foreslås, at disse hormoner er opstået fra den samme forløber i evolutionsprocessen. γ-LPG har en svagere lipotropisk aktivitet end beta-LPG.
Melanocytstimulerende hormon
Dette hormon, syntetiseret i hypofysens mellemlap, stimulerer biosyntesen af hudpigmentet melanin i dets biologiske funktion og fremmer en forøgelse af størrelsen og antallet af pigmentceller (melanocytter) i paddernes hud. Disse egenskaber ved MSH anvendes i biologisk testning af hormonet. Der findes to typer af hormonet: alfa- og beta-MSH. Det er blevet vist, at alfa-MSH ikke har artsspecificitet og har den samme kemiske struktur hos alle pattedyr. Dets molekyle er en peptidkæde bestående af 13 aminosyrerester. Beta-MSH har derimod artsspecificitet, og dets struktur varierer hos forskellige dyr. Hos de fleste pattedyr består beta-MSH-molekylet af 18 aminosyrerester, og kun hos mennesker er det forlænget fra aminosyreenden med fire aminosyrerester. Det skal bemærkes, at alfa-MSH har en vis adrenokortikotrop aktivitet, og dets effekt på dyrs og menneskers adfærd er nu blevet bevist.
Oxytocin og vasopressin
Vasopressin og oxytocin, som syntetiseres i hypothalamus, akkumuleres i hypofysens baglap: vasopressin i neuronerne i nucleus supraopticus og oxytocin i nucleus paraventrikularis. Derefter overføres de til hypofysen. Det skal understreges, at forløberen for vasopressinhormonet først syntetiseres i hypothalamus. Samtidig produceres proteinet neurophysin type 1 og 2 der. Det første binder oxytocin, og det andet binder vasopressin. Disse komplekser migrerer i form af neurosekretoriske granuler i cytoplasmaet langs axonen og når hypofysens baglap, hvor nervefibrene ender i karvæggen, og indholdet af granulerne kommer ind i blodet. Vasopressin og oxytocin er de første hypofysehormoner med en fuldstændig etableret aminosyresekvens. I deres kemiske struktur er de nonapeptider med én disulfidbro.
De pågældende hormoner har forskellige biologiske effekter: de stimulerer transporten af vand og salte gennem membraner, har en vasopressoreffekt, forstærker sammentrækninger af livmoderens glatte muskulatur under fødslen og øger udskillelsen fra brystkirtlerne. Det skal bemærkes, at vasopressin har en højere antidiuretisk aktivitet end oxytocin, mens sidstnævnte har en stærkere effekt på livmoderen og brystkirtlerne. Den primære regulator af vasopressinsekretion er vandforbrug; i nyretubuli binder det sig til receptorer i de cytoplasmatiske membraner med efterfølgende aktivering af enzymet adenylatcyklase i dem. Forskellige dele af molekylet er ansvarlige for at binde hormonet til receptoren og for den biologiske effekt.
Hypofysen, der er forbundet via hypothalamus med hele nervesystemet, forener det endokrine system til en funktionel helhed og bidrager til at sikre kroppens indre miljøs konstans (homeostase). Inden for det endokrine system udføres homeostatisk regulering baseret på princippet om feedback mellem hypofysens forlap og "mål"-kirtlerne (skjoldbruskkirtel, binyrebark, kønskirtler). Overskud af det hormon, der produceres af "mål"-kirtlen, hæmmer, og mangel på det stimulerer sekretionen og frigivelsen af det tilsvarende tropiske hormon. Hypothalamus er en del af feedbacksystemet. Det er i det, at receptorzoner, der er følsomme over for hormonerne i "mål"-kirtlerne, er placeret. Ved specifikt at binde sig til hormoner, der cirkulerer i blodet, og ændre responsen afhængigt af hormonkoncentrationen, overfører hypothalamiske receptorer deres effekt til de tilsvarende hypothalamiske centre, som koordinerer adenohypofysens arbejde og frigiver hypothalamiske adenohypofysiotrope hormoner. Hypothalamus bør således betragtes som en neuroendokrin hjerne.